

A. Pengertian membran plasma
Membran plasma adalah protoplasma yang menjaga isi sel dan memisahkan isi sel dengan lingkungan luar sel. Membran plasma bersifat selektif permeabel. Membran plasma memiliki sifat-sifat hidrofobik di bagian tengah dan sifat hidrofilik di permukaan luar maupun permukaan sistolik. Membran plasma terdiri dari senyawa-senyawa lipida, protein,karbohidrat,enzim dan ion. Komponen kimiawi yang terlihat secara morfologis adalah karbohidrat, protein, dan lipida.
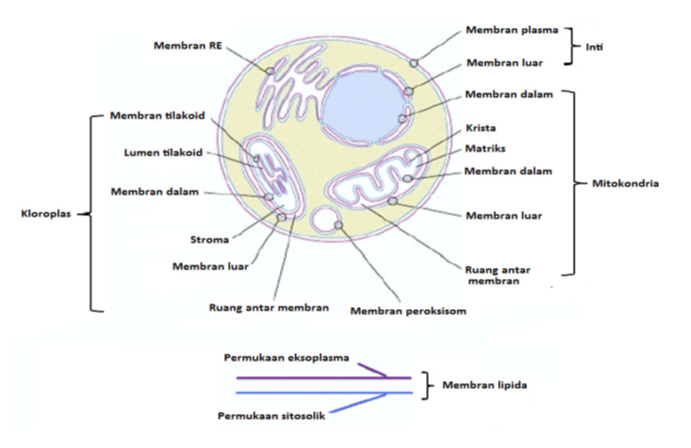
Gambar: letak-letak membran plasma
Fosfolipida merupakan lipida yang jumlahnya paling banyak dalam membran, membentuk dua lapisan yang disebut bilayer lipida, yang kemungkinan lapisan bagian atas diselipi protein dan bagian bawah tidak, atau keseluruhannya.Tipe Lipida pada membran plasama ada 3 macam yaitu Fosfogliserida ,Sfingolipida ,Kolesterol.
 |
Gambar: bagian-bagian
fosfolipid
|
Senyawa protein dan karbohidrat memberikan sifat hidrofilik.Struktur dasar selaput plasma ini sangat mendukung fungsinya sebagai pembatas lingkungan luar dari lingkungan dalam sel, dan lingkungan luar organela dari lingkungan dalamnya. Protein pada membran ada dua macam yaitu Protein ferifer (ekstrinsik) dan protein integral (intrinsik). Protein perifer umumnya tidak melekat dengan kuat pada membran dan mudah terlepas. Protein integral adalah protein yang berintegrasi dengan lipida.
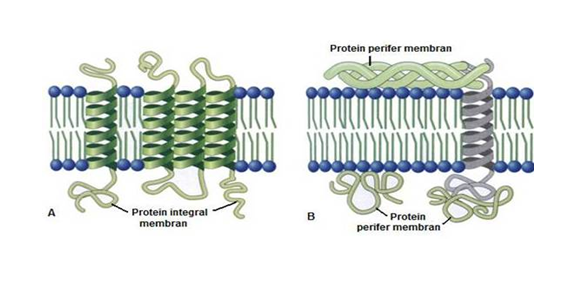 |
Gambar:jenis-jenis
protein
|
Karbohidrat yang menempel pada protein berupa glikoprotein dan karbohidrat yang menempel pada molekul lipida berupa glikolipida. Jenis karbohidrat yang menempel pada membran plasma tergantung pada fungsi membran plasma tersebut. Karbohidrat ada 3 jenis yang terdapat pada membran yaitu Oligosakarida,Glikoprotein:karbohidrat yang menempel pada protein,dan Glikolipid:karbohidrat yang menempel pada lipida.Karbohidrat tidak bisa menempel dengan protein atau lipida di bagian dalam kearah sistosilik karena karbohidrat akan dilisis.
Komposisi lipida, protein dan karbohidrat (pada glikoprotein dan glikolipida) bervariasi sesuai dengan macam selaputnya dan dapat berubah sesuai dengan tingkat perkembangan sel, umur, dan lingkungan. Keberadaan membran plasma sukar dilihat dengan mikroskop biasa dan keberdaan membran plasma sel dapat dibuktikan pada waktu sel mengalami plasmolisis atau sel yang dihilangkan dinding selnya. Dari segi fisiologis keberadaan membran plasma dapat dilihat dengan adanya permeabilitas selektif terhadap senyawa-senyawa tertentu. Di samping itu selaput plasma juga memperlihatka sifat-sifat yang dinamis antara lain pertumbuhan membran plasma, fragmentasi, diferensiasi dan perubahan struktur tiga dimensinya.
Komposisi masing-masing fraksi plasma terutama fraksi lipida sangat menenyukan kecairan selaput. Pengertian kecairan menyangkut ciri-ciri kekenyalan, kekentalan, atau kemudahan melakukan perubahan sifat fisiokemis untuk tetap dapat mempertahankan keutuhan fungsi membran plasma. Perubahan sifat fisiokemis itu terjadi dari keadaan seperti agar-agar ke keadaan lebih encer. Perubahan ke keadaan yang encer diikuti dengan peningkatan gerakan ikatan C-C molekul asam lemak pada poros sehingga terjadi susunan yang lebih acak. Perubahan ini menyerap tenaga panas yang besar.
Pada umumnya untuk berubah dari keadaan kental ke encer lipida dengan rantai karbon pendek atau rantai karbon dengan ikatan rangkap memerlukan tenaga transisi (tenaga panas) lebih rendah dibandingkan dengan asam lemak dengan rantai karbon panjang atau dengan ikatan-ikatan yang lebih sedikit (rantai C-C jenuh). Rantai karbon yang lebih pendek memerlukan luas permukaan yang lebih sedikit sehingga membuka peluang untuk adanya ikatan van der waals. Sebaliknya rantai karbon dengan ikatan rangkap menyebabkan konformasi berlipat-lipat sehingga memberikan bentuk lebih acak dan lebih sedikit ikatan van der waals yang terjadi serta lebih sedikit kemungkinan mengadakan interaksi dengan senyawa lipida lainnya.keadaan ini disebut dengan keadaan yang lebih encer. Dengan demikian kebanyakan sel mampu melakukan adaptasi terhadap suhu lebih rendah dengan meningkatkan bagian kandungan asam lemak tidak jenuh pada selaput plasma sehingga keenceran selaput plasma tetap dipertahankan.
Di samping tingkat kejenuhan asam lemak kolesterol juga merupakan penentu keenceran membran plasma. Molekul kolesterol terlalu bersifat hidrofobik sehingga lebih sukar untuk membentuk suatu bentukan lembaran. Dengan demikian kolesterol membentuk suatu interkalarsi di antara molekul-molekul fosfolipida. Gugus hidroksil kolesterol berada dekat gugus hidrofilik fosfolipida sedangkan cincin steroid berinteraksi dengan rantai karbon asam lemak, sehingga molekul asam lemak menjadi tidak mudah bergerak. Hasil akhir pengaruh kolesterol terhadap keenceran selaput bervariasi tergantung macam dan komposisi fosfolipida. Dengan kata lain kolesterol mengurangi gerakan asam lemak di dekat permukaan selaput plasma tetapi menyebabkan pemekaran ujung rantai karbon asam lemak atau mengurangi kemungkinan terjadinya ikatan antara rantai karbon asam lemak. Dengan demikian sebagai contoh selaput sitoplasmik pada organisme eukariotik yang mengandung lebih banyak kolesterol, selaputnya cenderung sulit berubah dari keadaan encer ke keadaan kental pada suhu sekitar 30 derajat C. hal ini diperlukan apabila suhu lingkungan lebih rendah. Struktur molekul penyusun selaput plasma diuraikan di bagian lain dalam.
Selain sebagai pembatas selaput plasma memiliki fungsi:
1. Mengatur permeabilitas terhadap senyawa-senyawa atau ion-ion yang melewatinya permeabilitas ini terutama diatur oleh protein integral.
2. Protein selaput yang berfungsi sebagai protein pengenal atau sebagai reseptor molekul-molekul khusus (hormon, antigen, metabolit) dan agensia khas (bakteri dan virus)
3. Protein selaput berfungsi sebagai enzim khusus misalnya pada selaput mitokondria, kloroplas, retikulum endoplasma, aparatus golgi selaput sel dan lain-lain.
4. Selaput sebagai kelompokan molekul juga berfungsi sebagai reseptor tehadap perubahan lingkungan seperti perubahan suhu, macam dan intensitas cahaya.
B. Pekembangan Model Selaput Plasma
1. Model Danielli-Davson
Danielli dan davson mendasarkan struktur selaput plasma melalui penelitian-penelitian fisiko-kimia yaitu dengan mebnadingkan tegangan permukaan yang terjadiantara tetes minyak pada ekstrak selaput plasma dan air. Tegangan permukaan yang terjadi antara tetes minyak dengan eksrak sel ternyata lebih rendah dibandingkan dengan tegangan permukaan yang terjadi antara tetes minyak dengan air. Hal ini menunjukkan adanya zat tertentu pada ekstrak sel yang mampu menurunkan tegangan permukaan pada batas kedua fasa. Pengamatan ini kemudian menghasilkan model selaput plasma yang disebut model Danielli-Harvey dan disempurnakan lagi sebagai model Danielli-davson. Beberapa kelemahan kedua model tersebut adalah tentang tidak adanya ketebalan selaput yang jelas apabila selaput plasma mengalami hidratasi.
Pengamatan dengan mikroskop elektron terhadap myelin dan beberapa selaput sel lainnya, menunjukkan gambaran dua garis sejajar seperti rel kereta api. Robertson mengusulkan suatu konsep struktural yang baru yaitu konsep selaput kesatuan. Dalam konsep tersebut digambarkan lapisan lipida sebagai dua lembaran molekul lipida disebut dwilembar lipida tang bagian hidrofiliknya bersinggungan dengan lapisan proteinberkonfigurasi memanjang atau berkonfigurasi B. Dengan model ini dapat diduga bahwa lapisan lipida membentuk ketebalan 3, 5 nm dan lapisan molekul protein setebal 2, 0 nm. Dengan demikian model ini dapat menjelaskan ketebalan selaput plasma yang umumnya mencapai 7, 5 nm.
 |
Gambar: membran plasma menurut Danielli-davson
|
2. S.J. Singer dan G. Nicolsan (1972)
mengemukakan bahwa protein membran itu terdispersi dan secara individual disisipkan ke dalam bilayer fosfolipida , dan hanya daerah-daerah hidrofiliknya yang menonjol. Menurut model ini, membran merupakan mosaik molekul protein yang terapung pada bilayer fluida yang terdiri dari fosfolipida-fosfolipida, yang dikenal dengan istilah membran model mosaik fluida.

3. E.Gorter dan F.Grandel (1925),
berpikir bahwa membran sel sebenarnya harus berupa bilayer fosfolipida yang terdiri dari dua lapisan moleku
C. Komposisi fraksi lipida membran plasma
Membran plasma terutama disusun dari molekul-molekul lipida dan protein. Kedua jenis molekul tersebut dapat mengalami glukosilasi menjadi glikolipida dan glikoprotein. Macam, panjang rantai dan pola ikatan glikosil sangat menentukan sifat dan fungsi membran plasma. Dengan demikian selaput plasma setiap organel dan sel memiliki keanekaragaman komposisi penyusunnya. Sebagian besar fraksi lipida merupakan lemak dwiasilgliserol, triasilgliserol, fosfilipid, dan glikolipida. Senyawa-senyawa asil gliserol disusun oleh senyawa gliserol, fosfogliserol yang berikatan ester dengan asam-asam lemak. Macam senyawa asam lemak sangat menemukan keenceran selaput plasma.
Senyawa-senyawa asam lemak kemudian dapat berikatan ester dengan gliserol menjadi mono, di, triasilgliserol. Senyawa=senyawa asam lemak dapat pula mengadakan ikatan ester dengan gliserol PO4 menjadi asam fosfatidat.
Asam-asam lemak dibedakan dan digolongkan berdasarkan jumlah C pada rantai –C, jumlah ikatan rangkap dan macam gugus reaktif yang terikat pada rantai C. panjang pendeknya rantai C dan jumlah ikatan rangkap sangat menentukan keenceran senyawa lemak. Dengan demikian senywa-senyawa gliserida sebagai penyusun selaput plasma dapat berpengaruh pada keenceran selaput plasma. Selain keanekaragaman asam lemak berdasarkan jumlah C dan ikatan rangkapnya, pada selaput plasma dijumpai pula keanekaragaman macam dan kadar fraksi lemak pada satu macam sel dan keanekaragaman komposisi berdasarkan jenis organela pada sel yang berbeda.
Dengan demikian sfingomielin dan fosfatidil kolin merupakan fosfolipida kha yang terdapat pada sisi luar, sedangkan fosfatidil etanolamin dan fosfatidil sering merupakan fosfolipida khas yang terdapat pada sisi sitoplasmik.
D. Komponen fraksi protein membran plasma
Protein membran plasma mempunyai fungsi yang sangat luas antara lain fungsi sebagai pembawa senyawa yang melewati membran plasma, menerima isyarat (signal) hormonal dan meneruskan isyarat tersebut ke bagian sel sendiri atau ke sel lainnya. Protein membran plasma juga berfungsi sebagai pangkal pengikat komponen-komponen sitoskeleton dengan senyawa-senyawa ekstraseluler. Molekul-molekul protein permukaan luar memberikan ciri-ciri individual tiap sel dan macam protein dapat berubah sesuai dengan diferensiasi sel. Protein-protein tersebut banyak yang berfungsi sebagai enzim (biokatalisator), terutama pada selaput mitokondria, kloroplas, retikulum endoplasma. Sebagai contoh, senyawa-senyawa fosfolipida selaput plasma disintesis oleh enzim-enzim selaput retikulum endoplasma.
Kedudukan dan orientasi protein dalam membran plasma bervariasi sesuai dengan selaput, macam sel dan jaringan. Orientasi molekul protein selaput dapat bersifat integral, porifer atau terbenam dalam bagian hidrofobik. Walaupun demikian beberapa orientasi ekstrem terlihat pada beberapa jenis sel antara lain: glikoforin pada selaput eritrosit merupakan suatu protein selaput yang hanya membentang sekali pada ketebalan selaput. Balteriorhodopsin terdapat pada selaput sel sejenis bakteri, merupakan protein selaput yang menembusselaput 7 kali. Protein-protein selaput dapat dilepaskan dari selaput plasma dengan menggunakan deterjen antara lain Triton-X, Octyl glucosyda, Sodium deoxhycholate, Cetylthrymhetillamonium bromide dan SDS.
E. Komponen fraksi karbohidrat membran plasma
Semua sel eukariotik memiliki molekul karbohidratpada permukaan luarnya. Senyawa-senyawa tersebut dapat berupa oligosakarida maupun polisakarida yang terikat secara kovalen pada protein selaput untuk membentuk glikoprotein atau berupa rantai oligosakarida yang terikat secara kovalen pada lipida selaput untuk membentuk glikolipida. Berat keseluruhan karbohidrat di dalam selaput berkisar antara 2 % sampai 10 % berat total selaput.
Membran plasma merupakan selaput yang asimetris. Molekul-molekul lipida lembaran luar membran plasma berbeda dengan molekul-molekul lipida yang terdapat di lembaran dalam. Demikian pula molekul polipeptida yang tersumbul dari kedua lembaran dwilapis lipida tersebut juga berbeda. Penyebaran karbohidratnya pun juga asimetris. Rantai-rantai karbohidrat dari sebagian besar glikolipida, glikoprotein dan proteiglikan pada selaput plasma tidak pernah ada di permukaan sistolik.
Ada sembilan macam monosakarida yang terikat pada glikoprotein dan glikolipid dari selaput plasma. Sakarida-sakarida terikat pada glikoprotein maupun glikolipida dengan ikatan –N glikosidik atau ikatan –O glikosidik. Di dalam glikolipida rantai monosakarida atau oligosakarida dihubungkan dengan ikatan – O glikosidik pada gugus hidroksil primer dari sfingosin. Glikoprotein dibentuk oleh ikatan kovalen residu gula pada salah satu residu-residu asam amoni dari rantai polipeptida. Asam amino tersebut adalah asparagin, serin, threonin, hidroksilisin, dan hidroksiprolin.
Salah satu cara yang tepat untuk menunjukkan keberadaan dan macam residu gula pada membran plasma yaitu teknik ikatan lektin. Lektin ini merupakan protein yang berasal dari bebrapa macam organisme, hewan maupun tumbuhan. Protein ini memiliki tempat untuk mengenali gula tertentu. Mengingat lektin dapat berikatan dengan glikoprotein maupun glikolipida selaput plasma, maka saat ini lektin digunakan sebagi alat penentu lokasi dan sarana untuk mengisolasi molekul-molekul selaput plasmayang mengandung gula.
Istilah selubung sel atau glikokaliks pada sel eukariotik digunakan untuk menyebut daerah bagian luar membran sel yang banyak mengandung karbohidrat. Daerah ini dapat ditunjukkan dengan berbagai cara antara lain dengan merah rutenium dan penandaan dengan lektin. Selain karbohidrat yang terikat pada protein integral dan fosfolipida selaput sel, glikokaliks dapat pula mengandung glikoprotein yang disekresikan dan kemudian menempel pada permukaan luar membran sel.
Struktur Dasar Membran Plasma Mikroorganisme Termofilik
Fraksi lipida selaput plasma mikroorganisme termofilik
Lipida membran plasma organisasi termofilik memiliki asam lemak-asam lemak dengan rantai karbon lurus, lebih jenuh dan lebih panjang dibandingkan dengan asam lemak selaput mikroorganisme mesofilik. Makin tinggi suhu sering dijumpai gugus metil. Dengan asam lemak tersebut mikroorganisme termofilik dapat mempertahankan derajat keenceran membran sel dengan demikian mempertahankan fungsi membran plasma pada suhu tinggi. Baik mikroorganisme termofilik maupun mesofilik dapat mengatur komposisi membran plasma. Pengendalian komposisi selaput ini disebut sebagai adaptasi homoeviscous.
Pada umumnya asam lemak memiliki 14-20 atom C dalam bentuk lurus dengan satu gugus metil dalam bentuk iso (2) atau anteiso (3). Bentuk iso dan anteiso bukan komponen yang khas pada mikroorganisme termofilik. Kedua-duanya juga umum ditemui pada mikroorganisme mesofilik. Walaupun demikian pada mikroorganisme termofilik kandungan lemak dalam bentuk iso dan anteiso merupakan bentuk yang dominan seperti tang dijumpai pada mikroorganisme mesofilik, komponen asam lemak pada eubakteri termofilik dapat terikat dengansenyawa karbohidrat dan dapat berkatan ester pada gliserol membentuk diasilgliserol.
Hasil analisis menunjukkan bahwa suhu lingkungan yang tinggi diikuti dengan pemanjangan rantai karbon. Penelitian Ray, et.al. menunjukkan bahwa kenaikan lipida total terutama glikolipida terlihat apabila suhu pertumbuhan ditingkatkan. Gejala seperti ini juga ditemukan pada mikroorganisme mesofilik.
Senyawa-senyawa alkohol golongan 1,2-diol merupakan konstituen umum pada lapisan lilin tanaman dan zat sebum pada hewan. Gabungan senyawa ini tidak pernah merupakan bagian dari membran sel. Walaupun demikian pada bakteri termofilik aerobik ditemukan alkohol 1,2-diol sebagai pengganti gliserol.
Senyawa-senyawa diol utama terdiri dari 21 atom C dan 19 atomC. Senyawa-senyawa ini memiliki gugus apolar dan merupakan 79 % dari bagian apolar lipida selaput dan 11 % asam lemak.
Pada bakteri termofilik juga dijumpai senyawa-senyawa alkohol isopropanol. Bakteri-bakteri mesofilik dan termofilik dapat mengubah komposisi bagia apolar sebagai tanggapan terhadap suhu linkungan. Dengan cara ini bakteri-bakteri tersebut dapat mengendalikan kekentalan selaput. Bagian apolar archaebacteria selalu mengandung senyawa tersebut dalam variasi C20 atau C40.
Senyawa monoalkohol isopropanoid c20 merupakan fitanol yang jenuh, sedangkan rantai isopropanoid C40 terdapat dalam bentuk 1,1 bifitanil-diol dengan kedua ujung terminal mengandung OH. Bentuk bifitanil-diol setara dengan 2 fitanol dengan kondensasi kepala ke kepala. Bila senyawa ini terikat dengan gliserol melalui ikatan eter, kedua alkohol tersebut menghasilkan 2 bentuk eter gliserol pada archaebacteria yaitu fitanilgliserol-dieter dan dibifitanil-digliserol tetraeter.
Bentuk dibifitanil-digliserol tetraeter ini memberikan struktur selaput ekalapis lipida (monolayer lipid membrane).
Fraksi protein pada selaput plasma
Mikroorganisme termofilik
Protein merupakan satu diantara biomolekul yang memegang peranan paling penting untuk melangsungkan sistem hidup yaitu protein sebagai enzim, protein sebagai penyusun ribosoma dan protein sebagai komponen selaput plasma. Sebaliknya protein merupakan biomolekul yang sangat peka dengan suhu tinggi. Walaupun demikian selaput plasma lebih termostabil dibandingkan dengan protein sitosplasmik. Dengan demikian pada mikroorganisme termofilik kemampuan untuk hidup pada suhu tinggi sangat tergantung pada adanya kekhasan struktural molekul protein. Ada petunjuk bahwa termostabilitas protein banyak ditentuka oleh adanya senyawa-senyawa poliamin yang terutama mempunyai peran melindungi fungsi protein sintesis invivo pada suhu tinggi. Sebagai contoh spermidin yang banyak dijumpai pada kebnyakan organisme. Mempunyai pengaruh pada asosiasi subunit ribosoma pada percobaan invitro terutama pada suhu tinggi dan kandungan Mg yang rendah. Pada percobaan lain thermin suatu poliamin yang didapatkan pada thermus sangat memacu protein pada suhu tinggi melalui pemacuan asosiasi subunit ribosoma, mRNA dan amino asil tRNA.
Peranan membran plasma
Pada pembahasan telah dikemukakan bahwa selaput plasma mempunyai beberapa peranan yang sangat penting pada kehidupan sel. Yang perlu mendapat perhatian yaitu peranannya sebagai penyekat, pemilih, pemilah dan pengatur. Selain itu sebagai plasma juga berperan sebagai tempat terjadinya reaksi kimia, sarana komunikasi, penerima dan penerus informasi dan lain sebagainya.
membran plasma sebagi penyekat.
Berbeda dengan sel prokariota yang hanya memiliki satu kompartemen yang diselubungi oleh membran plasma, sel-sel eukariota memilik sejumlah kompartemen berselaput yang disebut organela. Setiap kompartemen mempunyai peranan berbeda-beda mereka masing-masing memiliki seperangkat enzim, molekul-molekul khusus dan sistem distribusi yang rumit. Dalam bagian ini diuraikan sekilas tentang kompartemen dari suatu sel dan hubungan antar sel kompartemen-kompartemen tersebut.
Semua sel eukariota memiliki organela membran perangkat dasar. Adanya membran yang membatasi setiap organela ini sangat penting, karena kesgiatan di dalam setiap organela dapat berjalan lancar tanpa gangguan dari organela lain, namun tetap ada hubungan kerja yang serasi. Setiap organela berselaput memiliki sifat dan kemampuan yang serupa dengan semua jenis. Volume seluruh organela yang terdapat di dalam sel hampir setengah volume sel.
Membran plasma sebagai pengatur permeabilitas
Membran plasma tidak hanya merupakan penyekat pasif tetapi juga merupakan saringan pemilah yang antara lain memelihara perbedaan kadar ion di sebelah menyebelahnya. Dwilapis lipida berperan sebagai penyekat intermeabel bagi molekul yang terlarut dalam air dan molekul yang bermuatan. Materi yang melewati membran dikelompokkan menjadi dua kellompok yaitu mikro molekul dan makro molekul.
Pengangkutan mikromolekul lewat membran plasma
Hasil penelitian tentang pengangkutan lewat selaput plasma membuktikan bahwa terdapat tiga mekanisme pengangkutan yaitu difusi sederhana, difusi dipermudah atau dipercepat dan pengangkutan aktif. Difusi merupakan suatu proses lewatnya bahan-bahan tertentu melewati suatu membran sebagai akibat perbedaan kosentrasi.
Difusi sederhana dan difusi dipermudah keduanya merupakan transpor menurun yang berarti materi berasal dari daerah dengan konsentrasi tinggi ke daerah berkonsentrasi rendah. tenaga yang digunakan untuk pengangkutan ini merupakan tenaga panas atau tenaga termal. Mengingat bahwa pada pengangkutan ini tidak menggunakan ATP maka transpor ini dinyatakan sebagai transpor pasif. Transpor mendaki yaitu bila pengangkutan melawan derajat elektrokimia atau konsentrasi selalu memerlukan ATP selular. Oleh karena itu disebut transpor aktif. Perlu diingat bahwa pada sel yang hidup kegiatan pengangkutan ini berlangsung secara terpadu dan bersamaan untuk memelihara homeostatis sel.
Difusi sederhana
Molekul-molekul yang dapat melewati membran plasma secara difusi sederhana sangat terbatas, karena membran plasma masih memiliki penghalangnya. Mikromolekul terutama jenis hidrofobik dapat melewati selaput plasma dengan mudah, sedangkan makromolekul atau jenis molekul yang dapat terionisasi sulit melewati selaput plasma. Perbedaan ini biasanya dihubungkan dengan besarnya daya larut substansi hidrofobik di dalam dwi lapis lipida membran plasma.
Kemampuan sel dapat memilah senyawa hidrofilik dengan berat molekul (BM) kecil dari senyawa yang BM-nya besar, seringkali akibat adanya saluran akuosa atau porus pada membran plasma. Terdapat dua jenis porus jenis pertama merupakan saluran akuosa yang menembus molekul protein integral atau diantara kelompokan molekul protein transmembran. Porus jenis kedua disebut porus statistik yang terbentuk secara acak pada selaput plasma dan menembus dwilapis lipida.
Difusi dipermudah
Senyawa yang melewati membran plasma dengan jalan melewati difusi dipermudah, juga dapat memerlukan keterlibatan ATP, seperti halnya difusi dipermudah. Namun gerakan senyawa dari luar ke dalam atau sebaliknya lebih cepat dari difusi sederhana. Hal ini disebabkan adanya protein pembawa yang mampu mempercepat pengangkutan. Berdasarkan pemikiran ini suatu membran plasma pasti memiliki sejumlah protein pembawa yang masing-masing yang mempunyai tempat khusus untuk sesuatu molekul yang dapat diangkut.
Molekul protein pembawa setelah mengikat senyawa yang akan dibawa segera memindahkan senyawa tersebut dari luar ke dalam atau sebaliknya dengan jalan berputar, berdifusi atau membentuk poros.

Transpor aktif
Pengangkutan senyawa melewati selaput plasma dengan melawan gradien berlangsung dengan sangat rumit. Mekanisme yang paling sederhana mirip dengan difusi dipermudah namun memerlukan ATP. Terdapat dua kategori transpor aktif, transpor aktif primer yaitu transpor yang melibatkan ATP atau aliran elektron, transpor aktif sekunder yaitu transpor yang tergantung pada kekuatan selaput atau gradien ion atau tenaga kimiosmotik.

Dua transpor ini saling berkaitan, dalam arti mekanisme transpor aktif primer menimbulkan suatu gradien yang memungkinkan terjadinya transpor aktif sekunder. Salah satu contoh transpor aktif adalah pemompaan ion Na dan Ka.
Konsentrasi ion Ka dipertahankan untuk selalu lebih tinggi daripada di luar sel, sebaliknya konsentrasi ion Na di dalam sel diusahakan selalu lebih rendah daripada di luar sel. Ion Ka dan ino Na dua-duanya dipompa melawan gradien konsentrasi dan pemompaan dapat nerlangsung akibat terjadinya hidrolisis ATP. Hidrolisis ATP terjadi karena adanya enzim ATPase yang terdapat pada selaput plasma. Pada selaput plasma utuh yang berada pada sel, natrium mengangtifkan pemompaan dan memacu kegiatan ATPase dari dalam sel saja, kalium bekerja dari lingkungan luar membran plasma.
0 Response to "Membran plasma"
Post a Comment
Terima Kasih Telah Berkunjung di Pustaka Pandani
Silahkan komentar anda,
Salam
Irfan Dani, S. Pd.Gr