
Pada
pembahasan terdahulu telah dikemukakan bahwa selaput (membran) plasma mempunyai
beberapa peranan yang sangat penting pada kehidupan sel. Yang perlu mendapat
perhatian yaitu peranannya sebagai penyekat, pemilih, pemilah, dan pengatur.
Selain itu, sebagai plasma juga berperan sebagai tempat terjadinya reaksi
kimia, sarana komunikasi, penerima dan penerus informasi, dan lain sebagainya.
1. Selaput
plasma sebagi penyekat.
Berbeda dengan sel prokariota yang
hanya memiliki satu kompartemen yang diselubungi oleh selaput plasma, sel-sel
eukariota memilik sejumlah kompartemen berselaput yang disebut organela. Setiap
kompartemen mempunyai peranan berbeda-beda
mereka masing-masing memiliki seperangkat enzim, molekul-molekul khusus
dan sistem distribusi yang rumit. Dalam bagian ini diuraikan sekilas tentang
kompartemen dari suatu sel dan hubungan antar kompartemen-kompartemen dalam sel tersebut.
Semua sel eukariota memiliki
organela selput perangkat dasar. Adanya selaput yang membatasi setiap organela
ini sangat penting, karena kesgiatan di dalam setiap organela dapat berjalan
lancar tanpa gangguan dari organela lain, namun tetap ada hubungan kerja yang
serasi. Setiap organela berselaput memiliki sifat dan kemampuan yang serupa
dengan semua jenis. Volume seluruh organela yang terdapat di dalam sel hampir
setengah volume sel.
Beberapa organel memiliki selaput
rangkap, yaitu mitokondria, plastid, dan nucleus, sedangkan organel-organel
lain berselaput tunggal dan ada yang tidak berselaput (Ribosoma). Nukleus
mengandung genom utama dan merupakan tempat terpenting untuk sintesis ADN dan ARN.
Cairan yang mengelilingi nukleus disebut sitoplasma yang terdiri dari sitosol
dan organel-organel lainnya. Sitosol merupakan tempat sintesis protein dan
metabolisme intermedia. Metabolisme ini mencakup perombakan molekul-molekul
mikro dan pembentukan molekul makro. Sebagian besar selaput plasma yang berada
di dalam sel digunakan sebagai penyekat ruang labirintin retikulum endoplasma
terhadap organel yang lainnya dan memisahkannya dari sitosol. Permukaan
sitosolik selaput RE sebagian besar penuh ditempeli ribosom yang terlibat dalam
sintesis protein integral maupun terlarut. Selain menyintesis protein RE juga menghasilkan lipid terutama fosfolipid
selaput. Apparatus golgi berupa tumpukan kompartemen beselaput yang berbentuk
pinggan yang disebut sisterna golgi. Kedalam
sisterna ini tertuang lipid dan protein yang disintesis RE. Mekanisme
masuknya hasil sintesis RE ke sisterna golgi dibicarakan kemudian. Kloropls dan
mitokondria merupkana organella penghasil tenaga ATP yang digunakan untuk
melangsungkan reaksi biosintetik. Organella yang berperan merombak dari luar
sel disebut lisosoma. Organella ini mengandung enzim hidrolase yang bekerja
pada lingkungan asam. Selain organel yang telah disebut di atas, terdapat kompartemen-kompartemen yang
berbentuk vesikuli dan berisi enzim-enzim oksidatif. Selain itu terdapat pula
vesikuli yang berperan sebagai pengangkut antar organella yang satu ke organel
yang lain, demikian pula antara organella dan selaput (membran) sel.
Untuk mengetahui keterkaitan antar
kompartemen ditinjau dari segi perkembangannya. Sel eukariotik pertama diduga
mirip dengan bakteri yang hanya memiliki selaput sel tetapi tidak memiliki
selaput sitoplasmik. Selaput plasma pada sel semacam itu melakukan semua tugas
selaput plasma termasuk pemompaan ion, sintesis ATP, maupun sintesis lipid. Sel
eukariotik masa kini luas permukaan selaput plasmanya mencapai 10-30 kali luas
permukaan selaput plasma bakteri, misalnya E. Coli.
Evolusi
selaput sitoplasmik berjalan seiring dengan pengkhususan fungsi selaput plasma.
Pada beberapa bakteri masa kini di selaput plasmanya terdapat beberapa bercak
khas, padanya dijumpai sekelompok protein selaput yang melaksanakan gabungan
fungsi yang berkaitan. Bercak-bercak khas ini menggambarkan organel primitif.
Pada bakteri fotosintetik tempat bercak tersebut berkembang membentuk lekukan
(invaginasi). Lekukan ini dapat sedimikan dalamny sehingga akhirnya terlepas
dari selaputt plasma asalnya dan terbentuklah vesikuli berselaput khusus untuk
fotosintesis. Terbentuknya mitokondria dan kloroplas diduga karena terjadi
penyelubungan bakteri oleh selain yang semula hidup secara simbiosis. Nukleus dan
RE diduga berasal dari pelekukan selaput plasma yang mengandun DNA atau ribosom
dari organisme eukariota awal.
Selaput
plasma sebagai pengatur permeabilitas
Selaput plasma tidak hanya merupakan
penyekat pasif tetapi juga merupakan saringan pemilah yang antara lain
memelihara perbedaan kadar ion di sebelah menyebelahnya. Dwilapis lipida
berperan sebagai penyekat intermeabel bagi molekul yang terlarut dalam air dan
molekul yang bermuatan. Materi yang melewati selaput dikelompokkan menjadi dua
kellompok yaitu mikro molekul dan makro molekul.
Pengangkutan
mikromolekul lewat selaput plasma
Hasil penelitian tentang
pengangkutan lewat selaput plasma membuktikan bahwa terdapat tiga mekanisme
pengangkutan yaitu difusi sederhana, difusi dipermudah atau dipercepat dan
pengangkutan aktif. Difusi sederhana dan difusi dipermudah keduanya merupakan
transpor menurun yang berarti materi berasal dari daerah dengan konsentrasi
tinggi ke daerah berkonsentrasi rendah. tenaga yang digunakan untuk
pengangkutan ini merupakan tenaga panas atau tenaga termal. Mengingat bahwa
pada pengangkutan ini tidak menggunakan ATP maka transpor ini dinyatakan
sebagai transpor pasif. Transpor mendaki yaitu bila pengangkutan melawan
derajat elektrokimia atau konsentrasi selalu memerlukan ATP selular. Oleh
karena itu disebut transpor aktif. Perlu diingat bahwa pada sel yang hidup
kegiatan pengangkutan ini berlangsung secara terpadu dan bersamaan untuk
memelihara homeostatis sel.
Difusi
sederhana
Molekul-molekul yang dapat melewati
selaput plasma dengan jalan difusi sederhana sangat terbatas jumlahnya dan
untuk inipun selaput plasma masih memiliki penghalangnya. Mikromolekul terutama
jenis hidrofobik dapat melewati selaput plasma dengan mudah sedangkan
makromolekul atau jenis molekul yang dapat terionisasi sulit melewati selaput
plasma. Perbedaan ini biasanya dihubungkan dengan besarnya daya larut substansi
hidrofobik di dalam dwi lapis lipida selaput plasma.
Kemampuan sel untuk dapat memilah
senyawa hidrofilik dengan BM (berat molekul) kecil dari senyawa yang BM-nya
besar, seringkali akibat adanya saluran akuosa atau porus pada selaput plasma
tersebut. Terdapat dua jenis porus jenis pertama merupakan saluran akuosa yang
menembus molekul protein integral atau diantara kelompokan molekul protein transmembran.
Porus jenis kedua disebut porus statistik yang terbentuk secara acak pada
selaput plasma dan menembus dwilapis lipida.
Difusi
dipermudah
Senyawa
yang melewati selaput plasma dengan jalan melewati difusi dipermudah, juga
dapat memerlukan keterlibatan ATP, seperti halnya difusi dipermudah. Namun
gerakan senyawa dari luar ke dalam atau sebaliknya lebih cepat dari difusi
sederhana. Hal ini disebabkan adanya protein pembawa yang mampu mempercepat
pengangkutan. Berdasarkan pemikiran ini suatu selaput plasma pasti memiliki
sejumlah protein pembawa yang masing-masing yang mempunyai tempat khusus untuk
sesuatu molekul yang dapat diangkut. Molekul protein pembawa setelah mengikat
senyawa yang akan dibawa segera memindahkan senyawa tersebut dari luar ke dalam
atau sebaliknya dengan jalan berputar, berdifusi atau membentuk poros.

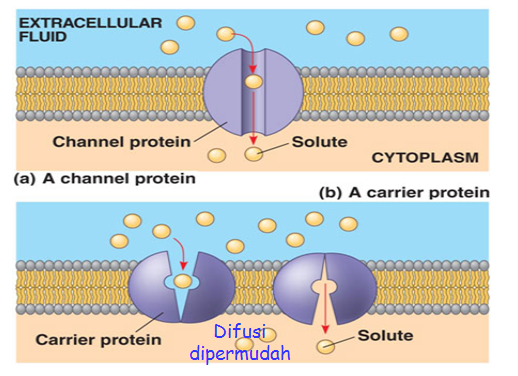 |
Gambar Difusi Melewati Membran
Ket.: (a) Difusi Sederhana, (b) Difusi yang dipermudah
|


Transpor
aktif
Pengangkutan
senyawa melewati selaput plasma dengan melawan gradien berlangsung dengan
sangat rumit. Mekanisme yang paling sederhana mirip dengan difusi dipermudah
namun memerlukan ATP. Terdapat dua kategori transpor aktif, transpor aktif
primer yaitu transpor yang melibatkan ATP atau aliran elektron, transpor aktif
sekunder yaitu transpor yang tergantung pada kekuatan selaput atau gradien ion
atau tenaga kimiosmotik. Dua transpor ini saling berkaitan, dalam arti
mekanisme transpor aktif primer menimbulkan suatu gradien yang memungkinkan
terjadinya transpor aktif sekunder. Salah satu contoh transpor aktif adalah
pemompaan ion Na dan Ka.
Konsentrasi
ion Ka dipertahankan untuk selalu lebih tinggi daripada di luar sel, sebaliknya
konsentrasi ion Na di dalam sel diusahakan selalu lebih rendah daripada di luar
sel. Ion Ka dan ino Na dua-duanya dipompa melawan gradien konsentrasi dan
pemompaan dapat nerlangsung akibat terjadinya hidrolisis ATP. Hidrolisis ATP
terjadi karena adanya enzim ATPase yang terdapat pada selaput plasma. Pada
selaput plasma utuh yang berada pada sel, natrium mengangtifkan pemompaan dan
memacu kegiatan ATPase dari dalam sel saja, kalium bekerja dari lingkungan luar
selaput plasma.

 |
Gambar
: Transpor Pasif dan Transpor Aktif
|
Transpor
aktif pada bakteria
Telah diketahui bahwa transpor aktif
dapat dikatakan dikemudikan oleh hidrolisis ATP gradien ion dan cahaya. Cara
transpor keempat umumnya terjadi pada bakteria berlangsung sebagai berikut:
molekul yang baru saja masuk dijerat sehingga tidak dapat keluar lagi lewat
jalan yang sama. Sebagai contoh: pada beberapa bakteria gula yang baru saja
melewati selaput segera difosforilasi menjadi gula fosfat. Mengingat bahwa
mereka mengion dan tidak dapat keluar maka gula fosfat menimbun di dalam sel.
Lagi pula gula yang masuk selalu difosforilasi akibatnya konsentrasi gula tanpa
fosfat di dalam sel menjadi sangat rendah. Dengan demikian gula dapat
terus-menerus masuk. Mekanisme fosforilasi pada
transpor ini sangat rumit dan teratur, serta melibatkan empat jenis
protein selaput dan fosfoenolpiruvat sebagai tenaga.
Pengangkutan
makromolekul lewat sitoplasma
Makromolekul seperti protein,
polinukleotida atau polisakarida tidak dapat lewat selaput plasma melalui
protein transmembran yang berperan sebagai pembawa. Namun sel tetap dapat
memasukkan dan mengeluarkan makromolekul-makromolekul tersebut. Pengangkutan
makromolekul ini sangat berbeda dengan pengangkutan mikromolekul. Pada
pengangkutan makromolekul terlihat pembentukan berurutan dan peleburan antara
selaput-selaput vesikuli dengan selaput sel. Terdapat iga cara pengangkutan
makromolekul endositosis yaitu pengangkutan makromolekul ke dalam sel.,
eksositosis yaitu pengeluaran makromolekul dari sel, yang ketiga dalah
pertunasan. Hal ini patut diperhatikan dalam proses-proses ini yaitu bahwa
makromolekul yang dimasukkan maupun dikeluarkan berada dalam vesikuli terpisah
dari makromolekul lainnya yang terlarut di dalam sitosol.
Terdapat tiga jenis endositosis:
•
Fagositosis
Pada fagositosis, sel menelan
suatu partikel dengan pseudopod yang membalut di sekeliling partikel tersebut
dan membungkusnya didalam kantong berlapis membran yang cukup besar unuk bisa
digolongkan sebagai vakuola.
•
Pinositosis
Sel meneguk tetesan fluida
eksraseluler dalam vesikula kecil. Karena salah satu atau seluruh zat terlarut
yang larut dalam tetesan tersebut dimasukkan ke dalam sel, pinositisis tidak bersifat spesifik.
Sebaliknya endositosis yang diperantarai reseptor sangat spesifik
•
Endositosis
yang diperantarai reseptor
Proses
pemasukan ciaran fluida ekstraseluler ke dalam sel yang bersifat spesifik. Yang
tertanam dalam membran adalah protein dengan tempat reseptor yang spesifik yang
dipaparkan ke fluida ekstraseluler. Reseptor yang utama dari membran adalah
karbohidrat berupa glikoprotein dan glikolipida





Oleh:
R I S T I O N O S O E G E N G
Diadop dari: ISSOEGIANTI S. M. R
R I S T I O N O S O E G E N G
Diadop dari: ISSOEGIANTI S. M. R
loading...
(function(){
var D=new Date(),d=document,b='body',ce='createElement',ac='appendChild',st='style',ds='display',n='none',gi='getElementById';
var i=d[ce]('iframe');i[st][ds]=n;d[gi]("M283033ScriptRootC165025")[ac](i);try{var iw=i.contentWindow.document;iw.open();iw.writeln("
0 Response to "Peranan Selaput (Membran) Plasma"
Post a Comment
Terima Kasih Telah Berkunjung di Pustaka Pandani
Silahkan komentar anda,
Salam
Irfan Dani, S. Pd.Gr